Archives
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
It should be noted that the long lifetime binding responsive
2019-07-02
It should be noted that the long-lifetime binding-responsive luminescence of ARC-1528 and ARC-1530 becomes evident only upon illumination with radiation at near-UV range where the excitation of ATB or ASB moiety is possible. At higher wavelength range (visible light), the fluorophore (5-TAMRA) can b
-
IAA sensitive Cl channels were shown
2019-07-02
IAA-94-sensitive Cl channels were shown to mediate cardioprotection due to IPC (Diaz et al., 1999) and cyclosporine A (Diaz et al., 2013). CLIC-like channel activity was also observed in cardiac mitoplast (Misak et al., 2013) but the molecular identity of these intracellular Cl channels and their pr
-
In conclusion the results obtained in this
2019-07-02
In conclusion, the results obtained in this study and those recently reported by our group (Abdel-Samad et al., 2012) clearly indicate that a dialogue indeed exists between the systems of NPY and ET-1 at the level of human endocardial endothelial cells, since these cells, whether isolated from the r
-
TAK-242 EP receptor is known to mediate the protective
2019-07-02
EP4 receptor is known to mediate the protective effect of PGE2 on the respiratory, gastrointestinal and glomerular epithelial lining (Aoudjit et al, 2006, Kabashima et al, 2002, Schmidt et al, 2011). In this study, the expression of asEP4 in the gills was limited to the epithelial lining suggesting
-
br Conclusion br Experimental NMR
2019-07-02
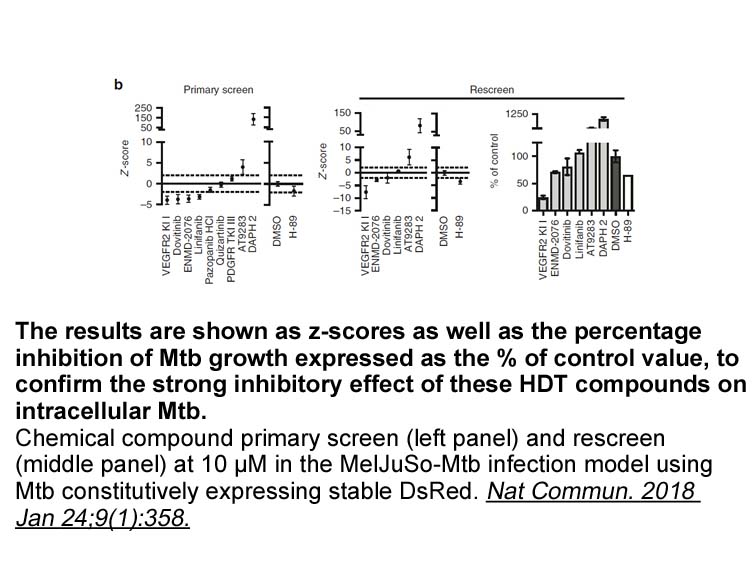
Conclusion Experimental NMR spectra were recorded on a Jeol spectrometer at either 270 or 400 MHz for 1H spectra and either 68 or 100 MHz for 13C spectra. All chemical shifts are quoted in ppm relative to tetramethylsilane. In the assignment of signals the abbreviation DHN is used for 2,3-dihy
-
MC 1568 structure The importance of EBI for B cell function
2019-07-02
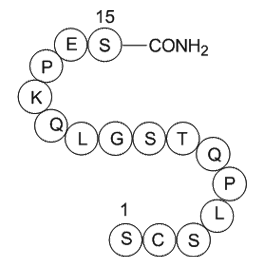
The importance of EBI2 for B cell function was first suggested by the dramatic upregulation of this receptor in EBV-transformed MC 1568 structure and further inferred from its regulation in activated and GC B cells (Birkenbach et al., 1993, Glynne et al., 2000, Shaffer et al., 2000). An involvement
-
Moving forward the new mathematical model opens doors
2019-07-02
Moving forward, the new mathematical model opens doors for understanding the SCF network, where activity of a component is blunted through mutation or altered in expression in diseased states, and during therapeutic intervention. Computational modeling could reveal underappreciated secondary or tert
-
br Deregulation of CDK Many neurotoxic or stress conditions
2019-07-01
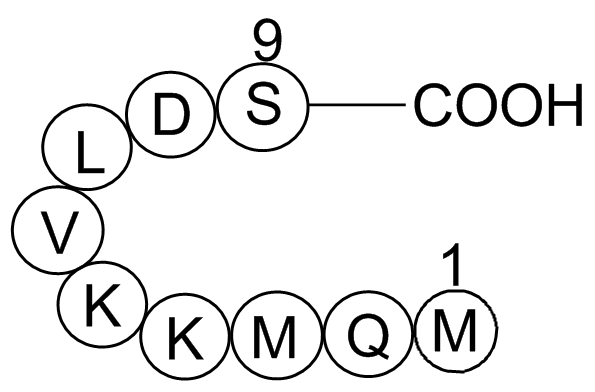
Deregulation of CDK5 Many neurotoxic or stress conditions like ischemic AS1842856 weight damage, oxidative stress, mitochondrial dysfunctions, excitotoxicity, Aβ exposure lead to disruption of the tight regulation of Cdk5 (De Strooper et al., 2010). Calpain-mediated cleavage of p35 to p25 is act
-
IL can induce phagocytosis cytoskeleton rearrangement gene e
2019-07-01
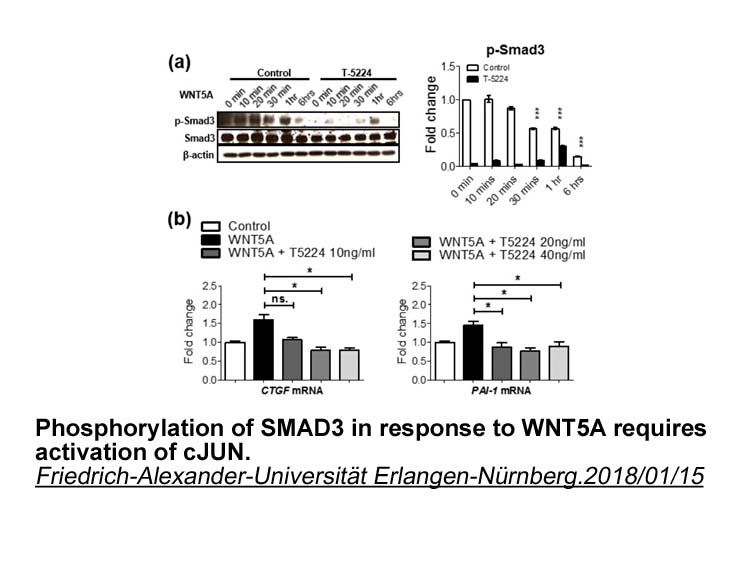
IL-15 can induce phagocytosis, cytoskeleton rearrangement, gene expression, de novo protein synthesis and can delay apoptosis in human neutrophils [15]. Production of chemokines, cytokines and natural inhibitors is increased in IL-15 induced neutrophils, including CXCL8 (IL-8) [16], IL-1beta, IL-1RI
-
Another problem is that the indications of each drug differ
2019-07-01
Another problem is that the indications of each drug differ among Asia-Pacific countries, and it neurokinin receptor is difficult to provide recommendations that are consistent with the indications in each country. For example, in Japan, the 3 novel OACs are indicated for “the prevention of ischemi
-
Introduction Implantable cardioverter defibrillators ICD car
2019-07-01
Introduction Implantable cardioverter-defibrillators (ICD), cardiac resynchronization therapy defibrillators (CRT-D), and cardiac resynchronization therapy (CRT-P) are useful tools for improving the prognosis and/or sudden cardiac death event rate in patients with heart failure and/or fatal ventric
-
Currently we are unable to fully understand the diversity
2019-07-01
Currently, we are unable to fully understand the ML-090 of and distribution of the AT arising around the apex of the triangle of Koch. Koyama et al. closely studied the spatial and topographic variation in the distribution of this type of AT around the His-bundle [17]. As demonstrated in the presen
-
br Introduction Epidemiological studies have shown that
2019-07-01
Introduction Epidemiological studies have shown that viral infections such as hepatitis C virus (HCV) and hepatitis B virus (HBV) are important factors contributing to hepatocellular carcinoma (HCC) development. Chronic HCV leads to cirrhosis in approximately 20% of patients over a 10–20 year per
-
GSK1016790A br Conclusion br Conflict of interest br Introdu
2019-07-01
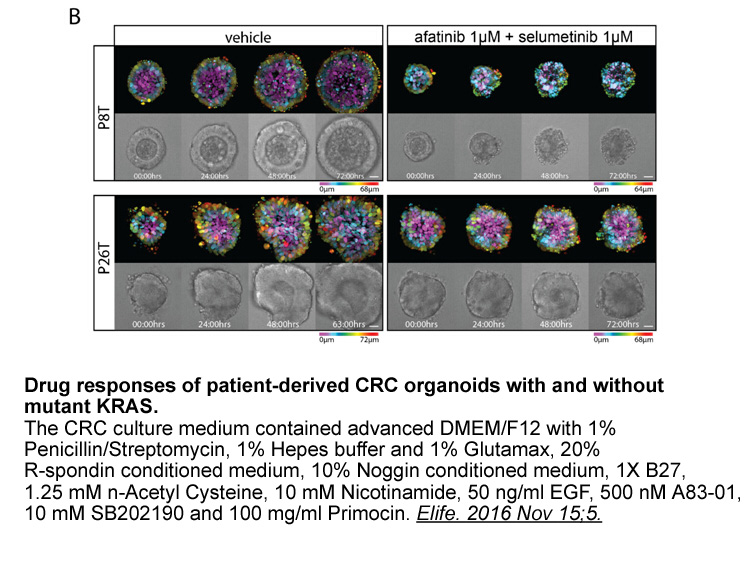
Conclusion Conflict of interest Introduction Future challenges and conclusion In recent decades several anticancer drugs have been approved for the management and treatment of colorectal cancer. These drugs do not represent a revolution in the treatment of CRC. However, 5-FU has continue
-
In this section we will try and address many of
2019-07-01
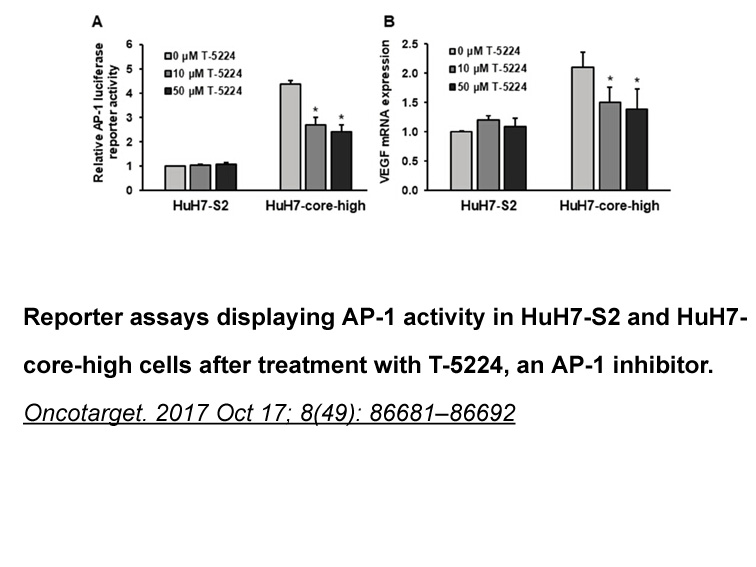
In this section we will try and address many of the frequently asked questions around preventive and prophylactic measures for patients either starting or already established on bone-targeted therapy (Figs. 2 and 3). In view of the difficulties in treating ONJ, preventive strategies would seem to ma
16666 records 1054/1112 page Previous Next First page 上5页 10511052105310541055 下5页 Last page